

Introduction
If you were to approach most people who possess a familiarity with paleontology, and ask them about what they know about raptor dinosaurs and how they lived their lives, these people would almost certainly tell you the following four things: they used their toe claws to kill their prey, they hunted in packs, they were fast, and they were smart. These four statements have been taken for granted for decades, but just how true are they? Were raptors really the super-predators of the Mesozoic world, or is it all Hollywood hype?
Killing Claws
Of course, the one anatomical feature that make raptors so easily recognizable are their enlarged claws on the second toe of their feet. These monstrous honking talons are often twice as big or even larger than the other main toes of the foot, and are often strongly curved into a hook. Restorations of raptor feet frequently show these enlarged claws suspended off of the ground by being retracted backwards, thereby allowing the claw to stick straight up while the remaining two toes were used for walking; footprint trackways found in Asia, which have been ascribed to raptor dinosaurs, seem to bear this out (1). Based upon appearance alone, these “killing claws” were surely used for grabbing and ripping open huge gashes in their victims. However, is that what they were really used for? Is there any way that you could prove that they were anatomically and physiologically capable of doing this?
It has been stated numerous times that raptors used these enlarged claws to kill their prey, with much academic speculation given as to just exactly how they went about doing it. The most common explanation which has been made states that the predator would physically grab or even jump onto the prey animal, gripping it with its large hand claws. Once anchored onto the prey, it would begin furiously kicking with its feet, making numerous deep gashes with their retractable switchblade-like killing claws, causing excessive blood loss and eventually death. This would enable small raptors to kill prey which was much larger than themselves.
One analog that we can perhaps use for how raptors used their killing claws is…the common house cat. Cats will sometimes grab things tightly with their arms, hugging it essentially, and then kick furiously with the claws on their feet. However, “kicking” doesn’t really accurately describe what’s happening here – they’re not punching things with their feet. Instead, what the cat is doing is using its exposed toe claws to latch onto whatever it has grabbed, and then sharply pull its leg backwards, using its foot claws to rake long scratches into whatever prey it has caught. Cats often do this repeatedly, not just once, to inflict as much damage with their toe claws as possible. Please watch the two videos below which demonstrate this behavior.
YouTube. Pomu & Shima. “ENG) My sister’s work won’t progress because Shima acts sweet…” (March 20, 2020).
YouTube. Pomu & Shima. “ENG) The angry Pomu took it out on me unreasonably…” (June 6, 2020).
Perhaps raptor dinosaurs, especially the smaller ones like Dromaeosaurus and Velociraptor, did the same thing. Their hand claws were often just as big or perhaps only slightly smaller as the “killing claws” on their feet. We also know from examining the arms, wrists, and hands of raptor dinosaurs that their hands were permanently oriented with the palms facing towards each other (2), which, while limiting, would still make them well-suited to bear-hugging (or in this case cat-hugging) things with both hands. It’s possible that small raptors like Velociraptor used their hands like grappling hooks, latching onto whatever prey they caught, while the head lunged downwards and latched onto the struggling prey, and then cat-kicking its prey by ripping it up over and over again with its foot claws.
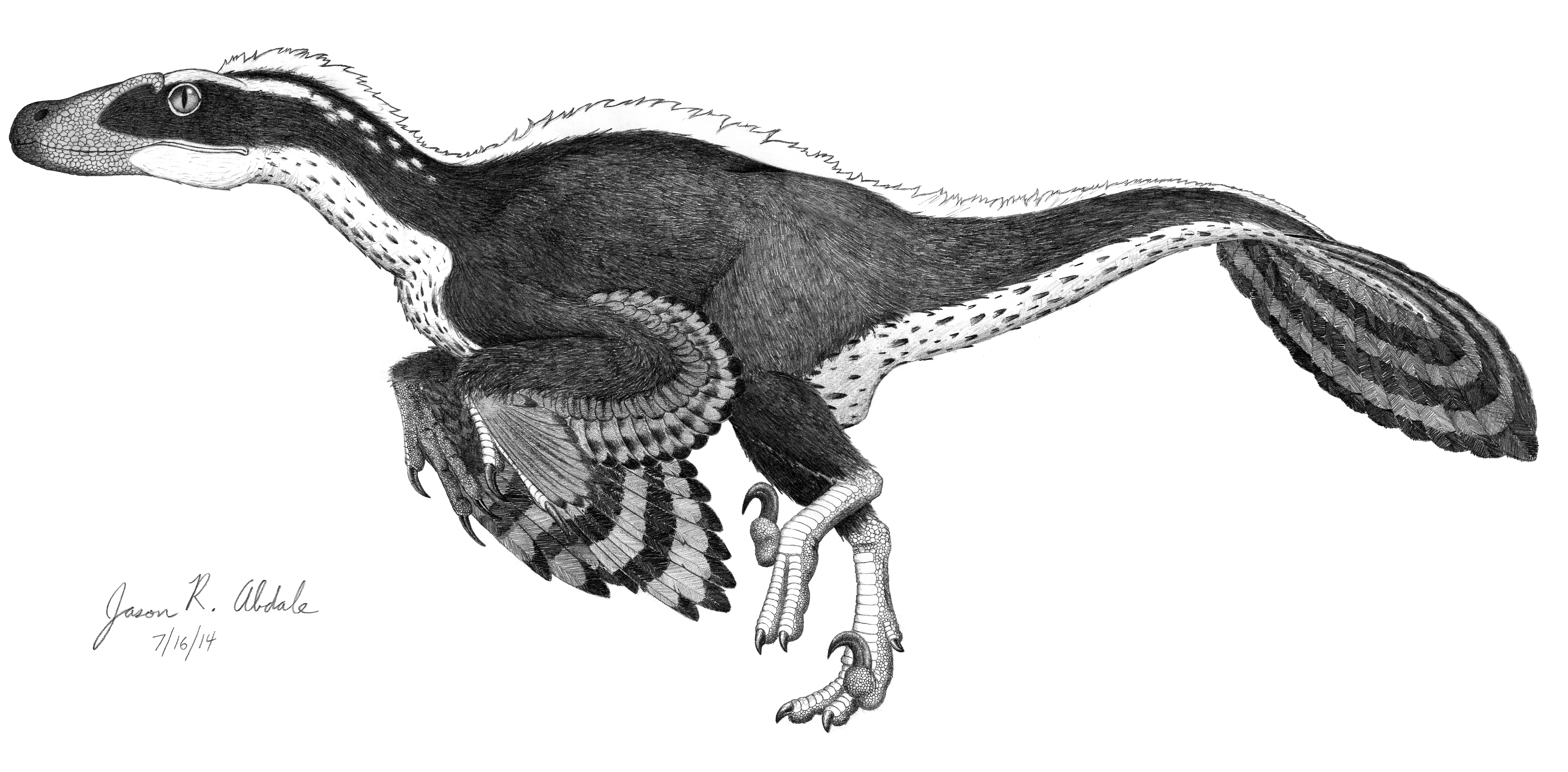
Acheroraptor. © Jason R. Abdale (July 16, 2014).
Other people are not so sure. If all raptors used their killing claws in the same fashion, then it would make sense that all raptor species would have killing claws which looked more-or-less identical. However, it’s clear from the fossil evidence that some raptor claws were more recurved than others. For example, the foot claws on Deinonychus are strongly curved into a hook, but the claws on Velociraptor are not and more closely resemble a farmer’s scythe. Some raptors had claws which were either larger or smaller than others in comparison to the other claws on their foot. For example, the “killing claw” of Dakotaraptor was downright gigantic, not only in terms of its sheer size but also the size of it in comparison to the other claws on its foot. On the opposite end of the spectrum, the Mongolian raptor Adasaurus had a “killing claw” which was the same size as its other foot claws (3).
It’s not just the claws which show variety, but the whole animal. There are many raptor species, and not all of them are built the same way. Broadly, raptors are divided into two families: the dromaeosaurids and the troodontids. The dromaeosaurids tend to be more muscular and stocky, while the troodontids have a much slimmer built. Dromaeosaurids like Deinonychus have a short compact metatarsus, which means that it could not run very fast (or at least couldn’t run for prolonged periods of time), but the foot would have possessed a very strong grip. By contrast, troodontids like Troodon and Saurornithoides tend to have a longer gracile metatarsus, which would make it more suitable for chasing after prey, but they would not have a strong grip (4).
One wonders if raptor claws were even physically capable of making those long ripping cuts, pulled through thick skin and tough muscle. Were the toes and feet of raptor dinosaurs strong enough to withstand such straining forces? Skeptics of the “slasher” hypothesis have pointed out that many of these claws are not well-designed for slashing, since they are not thin in cross-section and do not have a suitable blade-like cutting edge, and were instead used for gripping and pinning. In 2006, British paleontologist Philip Manning and his colleagues proposed that raptor toe claws were not properly designed to act as slashing weapons, and instead proposed that they acted like grappling hooks, securely anchoring the predator into the animal while it attacked with its teeth (5).
In order for its killing claw to be used in the way that we assume, it’s taken for granted that the muscles, tendons, and ligaments which operated this toe would have to be unusually strong compared to those which powered the other toes on the foot. A study conducted by Peter Bishop concluded that, under certain conditions, a Deinonychus killing claw could exert around 35 newtons of force, which is equivalent to around 7.86 pounds of force per square inch. However, this is the extreme top-end of the various results which were obtained through running numerous simulations of the physical strength of Deinonychus’ leg, and much lower forces of around 10-12 newtons (2.25 to 2.7 pounds per square inch) were far more common. Would that be sufficient enough to punch through the thick hide of a Tenontosaurus? I don’t think so. An analysis conducted in the same paper attempted to calculate the muscular strength of the leg, foot, and killing claw of Deinonychus, and concluded that the muscles, tendons, and ligaments which powered the enlarged foot claw were indeed formidably strong, but only under certain circumstances: “It was consistently found that more crouched whole-limb postures lead to higher claw forces being produced. This favours behavioural hypotheses that involve more flexed limb angles (such as grasping or restraining small to large prey at close quarters), and detracts from hypotheses involving an outstretched or strongly extended limb (such as slash kicking very large prey at a distance)” (6). In simple terms, it means that Deinonychus possessed a strong grip which would have been helpful in pinning down small prey animals, but it would have been extremely unlikely that Deinonychus used its talons for ripping open huge gashes in large prey animals – it simply did not have the strength to do such a thing.
The reference which Peter Bishop makes in this paper to hypotheses regarding raptors using their feet to grasp or restrain their prey comes from an earlier study which was made in 2011. During that year, Montana paleontologist Denver Fowler examined the “killing claws” of raptors and compared them with modern birds, and he noticed a strong similarity between the claws of raptor dinosaurs and the claws of birds-of-prey, specifically those of eagles and hawks. Several birds-of-prey (which are also known as “raptors”) have talons on the second claw of their foot which are noticeably larger than the talons on their remaining toes. However, these enlarged claws are not used for slashing or ripping – they’re used for grabbing. As further evidence of this, a re-examination of the foot structure of Deinonychus showed that the first toe on its foot had a unique orientation. In most reconstructions of theropod dinosaur feet, either in physical skeletons or models or in illustrations, the first toe, which is the one closest to the body and is the one that’s smallest in size, is oriented in the same way that the other toes are, curving backwards towards the posterior side of the foot. However, when Deinonychus’ foot was examined, the researchers found something odd – the dew claw was oriented sideways. This was not well-suited for walking, since the toe was still too short to touch the ground. However, it was well-suited for grabbing onto things. In fact, when the toes of the foot were flexed inwards, like the fingers of a human hand making a fist, they discovered something remarkable – the dewclaw acted almost like an opposable thumb. Deinonychus could grab prey with all of the toes on its foot, not just the main toes that it walked on (7).

Ventral view of Deinonychus foot (MOR 747) in flexion. Scale bar = 5 cm. (Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011)).
In Fowler’s view, raptor dinosaurs didn’t attack large animals, but rather went after prey which was the same size or smaller. According to his predation model, the raptor would run after and tackle the prey item, pinning it down with its enlarged claws. Then, it would lean down with its jaws to finish the job. Another scenario is that it would indeed jump on top of the animal, using its long pivoting tail and flapping its arms to maintain balance while the prey thrashed around, while its enlarged claws penetrated deeper and deeper into the prey animal. Perhaps, like modern-day hawks, it would even snap at and bite the prey while it was still moving around (which would explain why Deinonychus teeth are often found at kill sites), until the prey was gradually weakened by blood loss and perished (8).

Deinonychus. © Raul A. Ramos. Image used with permission.
This is not unheard of in today’s world. Large birds of prey like golden eagles sometimes attack juvenile deer, pouncing on their backs and gripping them tightly with their talons. Dr. Thomas R. Holtz Jr. of the University of Maryland also pointed out that such behavior, which is sometimes called “prey riding”, is seen in the Galapagos Hawk when it attacks marine iguanas. Eventually, the animal bleeds to death (9). The video below shows a Golden Eagle attacking a Pronghorn Antelope, gripping its back with its talons and ripping out small pieces of meat from its side while the animal is still walking around.
YouTube. Retire Cape Breton. “Eagle eating a deer alive” (December 31, 2018).
Pack Hunters
Ever since Deinonychus was revealed to the public in 1969, assertions have been made that it, and perhaps even all raptors, were social animals that lived in groups and hunted their prey in packs, but what’s the evidence to back this claim up?
John Ostrom had found the remains of four Deinonychus skeletons scattered around the remains of a single Tenontosaurus skeleton, which was five times larger. From examining the soil and rock layers, Ostrom concluded that these remains were not transported here by water and piled up in one spot, such as with Dinosaur National Monument which showcases the jumbled remains of hundreds of dinosaur skeletons that were washed on a riverbank following a devastating flash flood. Instead, the Deinonychus and Tenontosaurus died in-place, and they all appear to have died at around the same time. Ostrom hypothesized that these predators were working together as a pack to bring down an animal that was several times larger and stronger than they were. Unfortunately for them, the prey managed to kill four of them before it itself was killed (10).
The find that Ostrom made was not the only instance of Deinonychus and Tenontosaurus remains being found together. In fact, there are about 80 localities within the Cloverly Formation where Tenontosaurus fossils have been found, and of these, 35 of them also had Deinonychus teeth found nearby. It would appear that Tenontosaurus was the major prey item for Deinonychus. However, there seemed to be a size preference, as the majority of the Tenontosaurus skeletons which had associated Deinonychus teeth were juveniles which were only half-grown. A fully-grown 25 foot long Tenontosaurus would be extremely difficult for a single 10 foot long Deinonychus to take down, but a juvenile measuring only 10 to 15 feet long, most of it being tail, would be much more manageable (11).
In terms of the unfortunate Tenontosaurus which was found by John Ostrom and his dig team in 1964, it’s possible that the animal may have been killed by a single individual Deinonychus, and some other raptors who happened to be in the area were drawn towards the kill site by the scent of blood in the air and decided to help themselves to dinner. Or, it’s possible that, like modern-day crocodilians and Komodo Dragons, multiple individuals simply congregated in one spot and mob-killed the Tenontosaurus without any coordination or cooperation, resulting in a free-for-all as multiple Deinonychus tried to kill the animal at the same time (12). It’s also possible that this Tenontosaurus died through natural causes rather than being killed by the raptors, then the raptors happened upon the corpse, and a feeding frenzy broke out amongst the carnivores gathered there. The image that comes to mind is of a gaggle of feathered raptors squabbling amongst each other over a carcass like a noisy flock of buzzards. During this chaotic mayhem, the four Deinonychus which were found by Ostrom were killed by other members of their own kind in their contest over who could get their share of meat. Modern-day Komodo Dragons will sometimes kill and eat smaller Komodo Dragons which are trying to eat the same meat that the larger animal is trying to eat. As evidence of this, some of the bones on these Deinonychus skeletons exhibited bite marks – marks made by the teeth of other carnivorous dinosaurs. At another site in Montana where Deinonychus remains were uncovered, tooth marks were found on the animal’s hip, accompanied by a few spent teeth lying nearby. These teeth, and the associated bite marks, belonged to another Deinonychus. So, it appears that in addition to possible pack-hunting behavior, Deinonychus may have also attacked each other (claims made that this is evidence of cannibalism might be jumping to conclusions) (13).
Typically, paleontologists find three or four Deinonychus teeth associated with Tenontosaurus bones. However in 1992, a sub-adult Tenontosaurus skeleton measuring 14 feet long was discovered (collections ID code: MOR 682), and found close to it were eleven Deinonychus teeth of varying sizes. It would be very debilitating if any single Deinonychus lost eleven of its teeth in one instance, so the only conclusion is that these teeth came from the mouths of multiple Deinonychus which were feeding upon the corpse at the same time, which would be evidence of group hunting. The exact placement of these spent teeth has also been cited as evidence for pack-hunting behavior. All eleven teeth were found in the region of the animal’s underside, indicating that these teeth were all shed after the animal had been brought down; if the teeth had been shed while the Tenontosaurus was still up and kicking, then presumably these spent teeth would have been scattered all over the place. The fact that all of the teeth were found concentrated in one spot has been sited by some as evidence of cooperative group feeding on a freshly-made kill rather than a gaggle of individuals who just happened to be feeding on the same corpse (fresh or otherwise) at the same time (14).
However, I would like to counter this by saying that just because multiple individuals were feeding upon this dead animal, that doesn’t necessarily mean that they were all working together to bring it down and kill it. It must be observed that cooperative hunting amongst animals is extremely rare. Notable examples include killer whales, lions, hyenas, wolves, and African wild dogs – all of which are mammals. There are no modern-day reptiles which exhibit such behavior, despite claims made that some species of crocodilians do this (they don’t). As for birds, there is evidence that a few species of birds-of-prey hunt cooperatively in organized groups, such as the Red-Backed Hawk and the Harris Hawk, who also feed upon their prey animal based upon social hierarchy, with upper-ranking members of the group feeding first. However, Harris Hawks do not perform such behaviors habitually, but instead only operate in coordinated groups on occasion when the need calls for it. Observations of group behavior among Red-Backed Hawks also indicates that such behavior amongst them is “facultative” rather than “obligate”. Since dinosaurs are related to reptiles and birds, this puts the argument that raptors hunted cooperatively into a bit of a skeptical light (15). In terms of the sub-adult Tenontosaurus specimen designated as MOR 682, the location of the shed Deinonychus teeth around the belly of the sub-adult Tenontosaurus skeleton demonstrates that these individuals were feeding on the same area of the carcass, but it does not demonstrate that they were all part of a single social group. Carnivores will naturally attack the soft parts of a dead animal first since that’s where the most meat is and they don’t want to expend energy unnecessarily by crunching through bones. Fights could have broken out between the various carnivores over the choicest portions, which would explain the bite marks and claw marks which have been on Deinonychus bones which were likely made by other members of its kind as they scrapped with each other over getting their portion of meat.
Counter-claims such as these were put forward in a 2007 paper written by Brian Roach and Daniel Brinkman. They concluded that the evidence which was used by Ostrom to claim pack hunting behavior in Deinonychus was largely circumstantial, that there was no definitive evidence for pack hunting by Deinonychus, and therefore it was probably a solitary hunter. However, there was definitely evidence for violence amongst other members of its own species (click here for an article of my own which I wrote concerning the topic of intra-species combat amongst dinosaurs) (16).
There is one other line of evidence supporting pack hunting behavior – footprint trackways. A footprint trackway found in China, which was dated to the boundary of the Barremian and Aptian Stages of the Cretaceous Period (about 125 million years ago), was attributed to have been made by a group of raptor dinosaurs. These footprints belonged to multiple individuals, they were all moving in the same direction, and they all appear to have been made more-or-less at the same time. Some scientists have pointed to this as evidence that raptors lived and travelled in groups. In a 2008 article which was written about these footprints, the authors state “The tracks do not overlap, footprint size is almost uniform between trackways, and ‘intertrackway’ spacing (Lockley 1989) is regular and less than a single stride length, all of which strongly suggest that they were registered by a group of six individuals moving together as a coherent group, regardless of possible physiographic ‘control’. Because the sedimentology indicates a relatively rapid emplacement and subsequent burial of the trackways, a scenario in which six similarly sized individuals independently generated parallel, regularly spaced, nonoverlapping trackways is highly unlikely. Whether such an aggregation of similarly sized individuals represents a common or rare (or perhaps seasonal) event cannot be determined from a single find, but it indicates that dromaeosaurids in fact engaged in some type of group behavior” (17).

A diagram of the fossil footprint trackways found at the Houzuoshan Dinosaur Park located in Junan County, Shandong Province, China. The arrows which are denoted as D1 through D6 indicate the footprint pathways made by raptor dinosaurs. Li et al (2008), “Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China”. Naturwissenschaften, volume 95, issue 3 (April 2008). Page 188.
However, let’s not be too hasty. While the evidence as they present it is compelling, I think that the researchers may have been jumping to conclusions. I must point out that many of these “trackways” (which are visible on the diagram seen above), which are demoted ad D1 through D4, are incomplete, consisting of just a single footprint. A single footprint is not a trackway, it’s just a footprint. Saying that none of the trackways overlap, while true in the strictest sense, is quite presumptuous, as the researchers would have no idea if these trackways overlapped based upon just a single footprint from each trackway.
As mentioned earlier, Komodo Dragons will be drawn to the site of a kill from miles away by detecting the scent of blood in the air, and soon a kill site will be swarming with several dozen lizards of varying sizes. Just because a footprint trackway was discovered showing several individuals moving in the same direction, that does not mean that they were all moving together in a single group, and it does not mean that each of these individual trackways were all made at exactly the same time. Minutes or hours may have passed between the times that each individual animal came that way (18).
Another point of evidence is food. In social animals like lions or wolves, both the juveniles and the adults partake of the same meals and, naturally, eat the same kinds of food. You know the old expression “You are what you eat”? Well, it’s true. The type of foods that you eat most frequently will leave chemical signatures inside your teeth and bones. Isotopic analysis of the teeth of juvenile and adult lions and wolves show an identical signature – that’s no surprise, considering that both the young and the old are feasting on the flesh of the same meal. However, when you look at the teeth of animals that don’t live in social groups, you’ll see that there’s a marked difference. The isotopic signatures inside the teeth of juveniles verses those of adults is different, because the smaller juvenile animals are eating different kinds of food than the adults. When an isotopic analysis was done on the teeth of Deinonychus (even though they are fossils, the chemical traces are still present), it was revealed that the teeth of juvenile Deinonychus possessed a different chemical signature compared to the teeth of the adults. The juveniles and adults were eating different kinds of food, and that does not happen if the juveniles are living and eating alongside the adults (19).
As much as I find the image of pack-hunting raptors running and leaping after their prey to be evocative and alluring, the evidence to support this image is rather flimsy. By contrast, there is a growing pile of evidence which suggests that raptors, and Deinonychus specifically, didn’t live in packs after all, and were instead solitary ambush hunters who would occasionally congregate in groups to feed on carcasses, often fighting amongst themselves over the meat.
Speed
One attribute which is commonly given to raptors is that they were fast, supposedly. John Ostrom claimed that Deinonychus and its relatives were fast and agile runners (20). In the 1993 movie Jurassic Park, the game warden Robert Muldoon remarks that the Velociraptors housed within the park were as fast as cheetahs, being able to run at speeds of 50 or 60 miles per hour on broad open ground. But were raptors really that fast?

Dakotaraptor. © Jason R. Abdale (May 26, 2018).
Studies of primitive dromaeosaurs like Sinornithosaurus and primitive troodontids like Sinovenator show that both of these originally were indeed adapted for running. However, as evolution progressed, the troodontids retained their slim cursorial build while the dromaeosaurids became more adapted to walking or short-burst sprints, and altered the design of their feet to favor gripping strength (21).
A physical feature associated with a fast runner is a short thigh, a long shin, and a long metatarsus. Based upon this design, we can conclude that the dromaeosaurid raptor Deinonychus simply wasn’t built for the super-athletic lifestyle which it is often attributed to possess. Its lower leg bones (the tibia and fibula) were only slightly longer than the femur which forms the upper leg. Its metatarsal bones, which form the “flat” of the foot between the ankle joint and the toes, were short and stocky. If Deinonychus was built for speed, it would have had a leg constructed more like that of an ostrich – a short femur, a long shin, and a long metatarsus. Dinosaurs which do exhibit such a morphology are, not surprisingly, the “ostrich mimic” dinosaurs like Ornithomimus, Struthiomimus, and Gallimimus – these animals were built for speed. By contrast, Deinonychus does not appear to have been a fast runner. Instead, it appears to have been a short-distance sprint predator, lunging forwards with a sudden burst of speed. Such attributes are well-suited for ambush hunters, but they are not suitable at all to pursuit predators who actively chase after prey and run it down. Deinonychus wasn’t a cheetah – it was a leopard (22).
YouTube. Timm Martin “Bald Eagle Walking” (January 6, 2013).
What about other raptors? Deinonychus might not have been especially speedy, but what about the other smaller members of its group? Could the real-life Velociraptor really run as fast as a cheetah? Highly unlikely. The dromaeosaurid raptors, as a whole, had legs which were well-suited for walking or short-distance sprints – the hallmark of ambush predators. Their feet were not well-designed for running. However, they were well designed for grabbing, and the various joints in the metatarsus and the toes show that the foot as a whole possessed adaptations for torsion resistance. In other words, the feet were adapted for grabbing onto things which were still moving, twisting, and thrashing around. By contrast, the troodontid raptors have legs and feet which are much better suited for running than their dromaeosaurid cousins (23).

Velociraptor. © Raul A. Ramos. Image used with permission.
Intelligence
“All our theories about raptor intelligence, what they were capable of, we weren’t even close! They were smart. They were smarter than dolphins or whales. They were even smarter than primates!” – Dr. Alan Grant, Jurassic Park III (2000)
John Ostrom speculated that Deinonychus and its kind possessed a certain degree of intelligence (24), but were raptors really the brainiest animals of the Mesozoic? Well, I suppose that all depends on what exactly your definition of “intelligence” is.
Brain size in itself does is not indicative of intelligence, but rather, the size of the brain in relation to the size of the body as well as how complex the brain’s structure is. For example, Tyrannosaurus has THE largest brain of any dinosaur in terms of sheer size. However, in terms of how the brain worked, that’s another story. A study of dinosaur brain size which was conducted in 2013 showed that T. rex’s intelligence level lay on the borderline between that of crocodilians and birds. This would make it more intelligent than the primitive crocodilian-brained carnosaurs like Allosaurus and Carcharodontosaurus, but it would still be less intelligent than the more advanced bird-like carnivores such as raptors (25).
The ratio of brain size compared to body size is known as the “encephalization quotient”. Humans have an encephalization quotient or E.Q. of 1/40, meaning that an average human’s brain is one-fortieth of their total body weight. By contrast, it’s estimated that some of the larger dinosaurs like the long-necked sauropods may have had an E.Q. of 1/200, which would make their intelligence…well…questionable. An examination of the cranial cavities of raptor skulls showed that raptor dinosaurs did indeed possess larger brains relative to body size compared to other meat-eating dinosaurs. Not only were raptor brains proportionally larger, but they were also more complexly constructed than their non-raptor ancestors. Raptors belonged to a group of theropod dinosaurs called “Maniraptora”, which was the most advanced theropod group. Maniraptoran theropods possessed more developed brains than non-maniraptorans. Earlier theropods like megalosaurs and carnosaurs possessed primitive crocodilian-like brains, whereas advanced theropods like ornithomimids, oviraptorosaurs, and especially raptors, which were more closely related to birds than to crocodiles, possessed more bird-like brains. However, while it’s true that the brain cases of raptors were larger in proportion with the rest of their body compared to other meat-eating dinosaurs like Allosaurus and Tyrannosaurus, raptors were assuredly not as smart as popular perceptions make them out to be (26).
Another contributing factor to the long-held belief that raptors were smart comes from inference. It was believed for a long time that all raptors were pack hunters. Pack hunting requires a great deal of cooperation and coordination, and to do that, you need heightened intelligence – ergo, raptors were smart. The problem with this claim is that, as described earlier, the notion of raptors being pack hunters was based largely upon circumstantial evidence. From this, an entire mythos grew around these animals, ascribing traits to them which they probably never had.
One of the raptor dinosaurs named Troodon has been regarded for quite some time as the smartest dinosaur since it had the largest brain in proportion to body size of any known dinosaur. Troodon’s brain was purportedly so large that it left impressions on the inside of its cranial cavity (27). In fact, during the 1980s, the paleontologist Dale Russell proposed that if dinosaurs hadn’t gone extinct, creatures like Troodon would have continued to evolve into a humanoid appearance, which was christened by the media as “the dinosauroid”. Professor Russell based his claim on the notion that, in his opinion, evolution seemed to favor certain physical forms, and that different groups of animals were evolving to change into those forms (28). It’s an out-dated Victorian idea which is strongly based upon “Whig history” and its obsession with human progress culminating in the perfection of society, which in their minds was embodied in the British Empire. 18th Century and 19th Century naturalists took these ideas and applied them onto biology and claimed that life was striving to reach an ideal form, which of course in their eyes, were humans, since human beings were regarded as the perfection of life on Earth as dictated in the Book of Genesis (29). Such sentiments were echoed in the 1932 movie The Island of Lost Souls, a film adaptation of H. G. Wells’ 1897 book The Island of Doctor Moreau, in which the sinister doctor claims “All animal life is tending toward the human form”. Despite these whiggish notions and Dale Russell’s musings, it is extremely unlikely that Troodon would have evolved into a human-like creature.
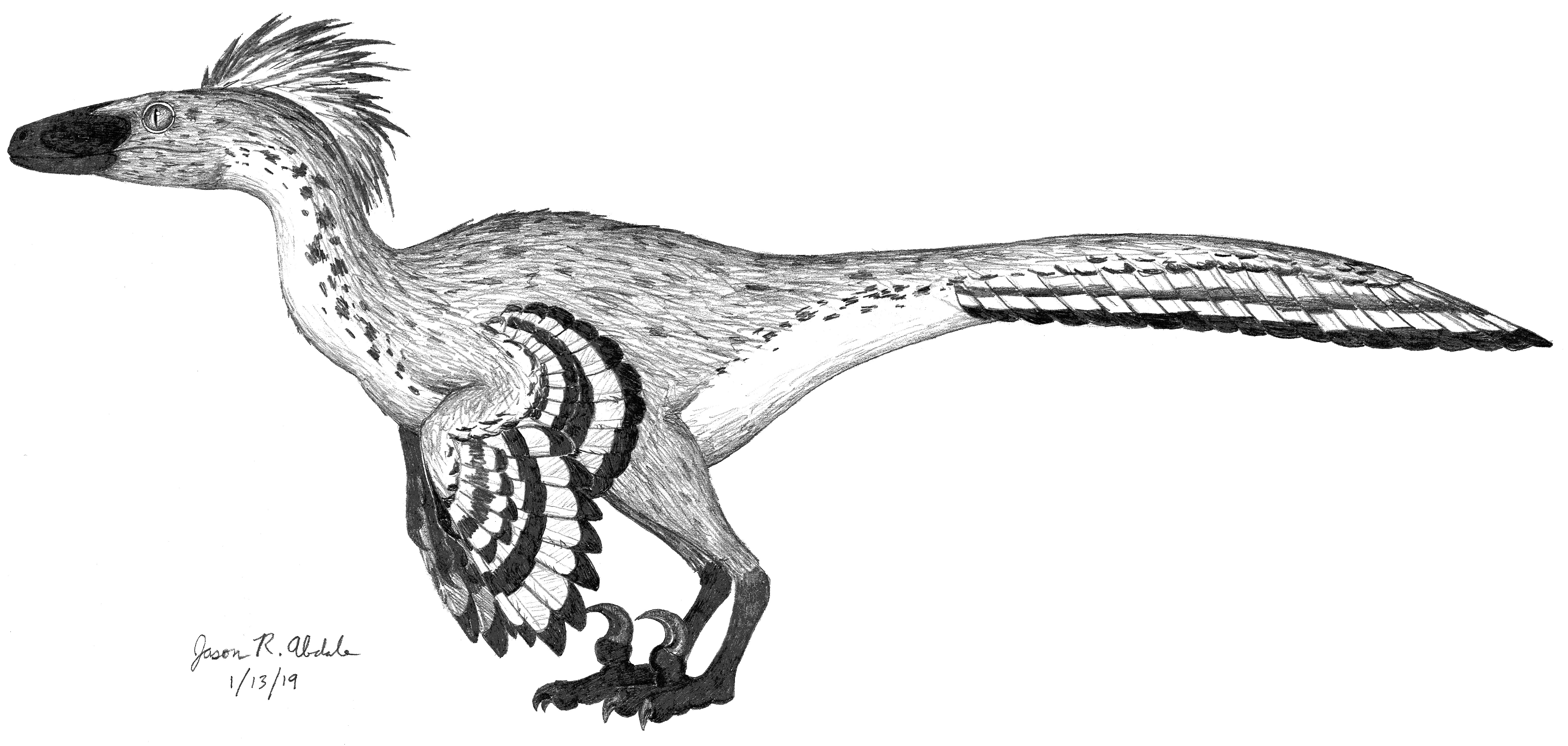
Troodon. © Jason R. Abdale (January 13, 2019).
However, it’s possible that Troodon’s intelligence has been grossly exaggerated (my apologies to all of the dead Troodons who may have been insulted by that statement). When Dale Russell plotted Troodon’s intelligence on a ratio curve comparing brain size to body size, he found that it ranked alongside primitive rodent-like mammals in terms of smarts. So, a Troodon would have been about as smart as a modern-day opossum, and it’s debatable just how smart these modern marsupials are – some studies have found that opossums proved unresponsive to performing even simple tasks, while other studies showed them to be just as smart as some breeds of dogs. The paleontologist Dr. Hans Larsson noted that the cerebral hemisphere, which forms the thinking part of the brain, is larger in proportion to overall brain size within Troodon than in any other dinosaur. Even so, he also stated that Troodon’s intelligence would have been comparable to that of a modern chicken (30), which would certainly be smarter than your average dinosaur, but still nowhere near as smart as the popular media has portrayed them to be.
Conclusion
For decades, scientists and the general public alike have affirmed that raptors were the super predators of the Mesozoic world. They were swift, smart, and deadly. They were dinosaurian wolves, roving the land in search of prey, living in tight-knit social groups, out-running, out-smarting, and out-gunning anything around.
However, based upon the evidence that we now have, it would appear that nearly all of the stereotypes about raptor dinosaurs which have been perpetuated since the late 1960s are wrong. Rather than living in packs, actively chasing after their prey, outsmarting it with their well-developed brains, and then bringing it down with several well-placed swipes of their killing claws, raptors were more likely to live their lives as prowling ambush-hunting loners rather than living and hunting in social groups, pouncing on their unsuspecting victims and tackling them to the ground like pro football players, and then savagely biting them over and over again. They lived their lives more like ground-hunting eagles or hawks rather than wolves.
All of this goes to show that it isn’t wise to take assumptions and treat them like unchallengeable dogma. Scientists always need to look at new evidence as it presents itself, and perhaps more importantly, they have to be willing to change their ideas based upon that new evidence. In doing so, we treat these creatures like actual flesh-and-blood animals, studying what they were truly like, rather than stubbornly clinging to our fanciful mythologized concepts of what we want them to be like. Our current ideas about raptors, which have already been substantially revised since the early 2000s, might prove to be just as erroneous as our previous notions about these animals were back in the 1970s. It is entirely possible that our knowledge about these animals will be altered even further with the discovery of even more evidence in the years to come.
Source citations
- Peter J. Bishop (2019), “Testing the function of dromaeosaurid (Dinosauria, Theropoda) ‘sickle claws’ through musculoskeletal modelling and optimization”. PeerJ, 7 (August 28, 2019): e7577.
- Alan D. Gishlick (2001), “The functional morphology of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight”. From New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John D. Ostrom, edited by Jacques Gauthier and L. F. Gall. New Haven: Peabody Museum of Natural History, 2001. Pages 301-318; Kenneth Carpenter (2002), “Forelimb biomechanics of nonavian theropod dinosaurs in predation”. Palaeobiodiversity and Palaeoenvironments, volume 82, issue 1. Pages 59-75.
- Mark Witton Blogspot. “Realistic raptors: pop-culture dromaeosaurs vs current science, part 1”, by Mark Witton (March 27, 2020); Peter J. Bishop (2019), “Testing the function of dromaeosaurid (Dinosauria, Theropoda) ‘sickle claws’ through musculoskeletal modelling and optimization”. PeerJ, 7 (August 28, 2019): e7577.
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011); “Deinonychus and Velociraptor used their killing claws to pin prey, like eagles and hawks”, by Ed Yong (December 14, 2011).
- Philip L. Manning, David Payne, John Pennicott, Paul M. Barrett, and Roland A. Ennos (2006). “Dinosaur killer claws or climbing crampons?”. Biology Letters, volume 2, issue 1 (March 22, 2006). Pages 110-112.
- Peter J. Bishop (2019), “Testing the function of dromaeosaurid (Dinosauria, Theropoda) ‘sickle claws’ through musculoskeletal modelling and optimization”. PeerJ, 7 (August 28, 2019): e7577.
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011).
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011); “Deinonychus and Velociraptor used their killing claws to pin prey, like eagles and hawks”, by Ed Yong (December 14, 2011).
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011); “Deinonychus and Velociraptor used their killing claws to pin prey, like eagles and hawks”, by Ed Yong (December 14, 2011).
- Dinosaur!, episode 3 – “The Tale of an Egg”. A&E, 1991; Natural History Magazine. “Day of the Deinos: Did predatory dinosaurs leave clues to their pack-hunting habits at kill sites?” by Desmond Maxwell (December 1999-January 2000); Beyond Bones – Blog of the Houston Museum of Natural Science. “Raptors – Group Hunters or Cannibals?”, by Robert Bakker (March 11, 2010).
- Brian T. Roach and Daniel L. Brinkman (2007), “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History, volume 48, issue 1 (April 2007). Page 111; Paleoworld – “Killer Raptors”; Natural History Magazine. “Day of the Deinos: Did predatory dinosaurs leave clues to their pack-hunting habits at kill sites?” by Desmond Maxwell (December 1999-January 2000).
- Brian T. Roach and Daniel L. Brinkman (2007), “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History, volume 48, issue 1 (April 2007). Pages 110-113; Fossil Crates. “Pack Hunting Raptors – What is the evidence?”, by Dr. Brian Curtice (May 21, 2020).
- Brian T. Roach and Daniel L. Brinkman (2007), “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History, volume 48, issue 1 (April 2007). Pages 110-118; Beyond Bones – Blog of the Houston Museum of Natural Science. “Raptors – Group Hunters or Cannibals?”, by Robert Bakker (March 11, 2010); Fossil Crates. “Pack Hunting Raptors – What is the evidence?”, by Dr. Brian Curtice (May 21, 2020).
- Paleoworld – “Killer Raptors”; Natural History Magazine. “Day of the Deinos: Did predatory dinosaurs leave clues to their pack-hunting habits at kill sites?” by Desmond Maxwell (December 1999-January 2000).
- James C. Bednarz (1988), “Cooperative Hunting in Harris’ Hawks (Parabuteo unicinctus)”. Science, volume 239, issue 4847 (March 25, 1988). Pages 1525-1527; Jennifer O. Coulson and Thomas D. Coulson (2013), “Reexamining Cooperative Hunting in Harris’s Hawk (Parabuteo unicinctus): Large Prey or Challenging Habitats?”. The Auk, volume 130, issue 3 (July 1, 2013). Pages 548-552; Sergio Alvarado Orellania and Ricardo A. Figueroa Rojas (2005), “Possible social foraging behavior in the Red-Backed Hawk (Buteo polyosoma)”. Ornitologia Neotropical, volume 16 (2005). Pages 271-275; Brian T. Roach and Daniel L. Brinkman (2007), “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History, volume 48, issue 1 (April 2007). Pages 103-110; Fossil Crates. “Pack Hunting Raptors – What is the evidence?”, by Dr. Brian Curtice (May 21, 2020).
- Brian T. Roach and Daniel L. Brinkman (2007), “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History, volume 48, issue 1 (April 2007). Pages 103-138.
- Rihui Li, Martin G. Lockley, Peter J. Makovicky, Masaki Matsukawa, Mark A. Norell, Jerald D. Harris, Mingwe Liu (2008), “Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China”. Naturwissenschaften, volume 95, issue 3 (April 2008). Pages 189-190.
- Fossil Crates. “Pack Hunting Raptors – What is the evidence?”, by Dr. Brian Curtice (May 21, 2020).
- J. A. Frederickson, M. H. Engel, and R. L. Cifelli (2020). “Ontogenetic dietary shifts in Deinonychus antirrhopus (Theropoda; Dromaeosauridae): Insights into the ecology and social behavior of raptorial dinosaurs through stable isotope analysis”. Palaeogeography, Palaeoclimatology, Palaeoecology, volume 552. (August 15, 2020); Fossil Crates. “Pack Hunting Raptors – What is the evidence?”, by Dr. Brian Curtice (May 21, 2020).
- Dinosaur!, episode 3 – “The Tale of an Egg”. A&E, 1991.
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011).
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011).
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011).
- Dinosaur!, episode 3 – “The Tale of an Egg”. A&E, 1991.
- Grant R. Hurlburt, Ryan C. Ridgely, and Lawrence M. Witmer (2013), “Relative Size of Brain and Cerebrum in Tyrannosaurid Dinosaurs: An Analysis using Brain-Endocast Quantitative Relationships in Extant Alligators”. From Tyrannosaurid Paleobiology, edited by J. Michael Parrish, Ralph E. Molnar, Philip J. Currie, and Eva B. Koppelhus. Bloomington: Indiana University Press, 2013. Page 135; Walking with Dinosaurs: The Ballad of Big Al. BBC, 2001; Dino Lab. The Discovery Channel, 2006.
- Dino Lab. The Discovery Channel, 2006.
- Paleoworld – “Troodon: Dinosaur Genius”.
- Dinosaurs, Dinosaurs, Dinosaurs, 1985; National Geographic. “Troodon sapiens?: Thoughts on the ‘Dinosauroid’”, by Riley Black (October 23, 2007).
- Dinosaur!, episode 1 – “The Tale of a Tooth”. A&E, 1991.
- Paleoworld – “Troodon: Dinosaur Genius”; Dino Lab. The Discovery Channel, 2006.
Bibliography
Articles:
- Bednarz, James C. (1988). “Cooperative Hunting in Harris’ Hawks (Parabuteo unicinctus)”. Science, volume 239, issue 4847 (March 25, 1988). Pages 1525-1527.
- Bishop, Peter J. (2019). “Testing the function of dromaeosaurid (Dinosauria, Theropoda) ‘sickle claws’ through musculoskeletal modelling and optimization”. PeerJ, 7 (August 28, 2019): e7577. https://peerj.com/articles/7577/.
- Carpenter, Kenneth (2002). “Forelimb biomechanics of nonavian theropod dinosaurs in predation”. Palaeobiodiversity and Palaeoenvironments, volume 82, issue 1. Pages 59-75. https://www.researchgate.net/publication/225366451_Forelimb_biomechanics_of_nonavian_theropod_dinosaurs_in_predation.
- Coulson, Jennifer O.; Coulson, Thomas D. (2013). “Reexamining Cooperative Hunting in Harris’s Hawk (Parabuteo unicinctus): Large Prey or Challenging Habitats?”. The Auk, volume 130, issue 3 (July 1, 2013). Pages 548-552. https://bioone.org/journals/the-auk/volume-130/issue-3/auk.2013.120063/Reexamining-Cooperative-Hunting-in-Harriss-Hawk-Parabuteo-unicinctus–Large/10.1525/auk.2013.120063.full.
- Fowler, Denver; Freedman, Elizabeth A.; Scannella, John B.; Kambic, Robert E. (2011). “The Predatory Ecology of Deinonychus and the Origin of Flapping in Birds”. PLoS ONE, volume 6, issue 12 (December 14, 2011). https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0028964.
- Frederickson, J. A.; Engel, M. H.; Cifelli, R. L. (2020). “Ontogenetic dietary shifts in Deinonychus antirrhopus (Theropoda; Dromaeosauridae): Insights into the ecology and social behavior of raptorial dinosaurs through stable isotope analysis”. Palaeogeography, Palaeoclimatology, Palaeoecology, volume 552. (August 15, 2020). https://par.nsf.gov/servlets/purl/10172318.
- Gishlick, Alan D. (2001). “The functional morphology of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight”. From New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John D. Ostrom, edited by Jacques Gauthier and L. F. Gall. New Haven: Peabody Museum of Natural History, 2001. Pages 301-318. https://www.researchgate.net/profile/Alan-Gishlick/publication/34256336_The_functional_morphology_of_the_forelimb_of_Deinonychus_antirrhopus_and_its_importance_for_the_origin_of_avian_fligth_microform/links/5665a00008ae418a786f1a42/The-functional-morphology-of-the-forelimb-of-Deinonychus-antirrhopus-and-its-importance-for-the-origin-of-avian-fligth-microform.pdf.
- Hurlburt, Grant R.; Ridgely, Ryan C.; Witmer, Lawrence M. (2013). “Relative Size of Brain and Cerebrum in Tyrannosaurid Dinosaurs: An Analysis using Brain-Endocast Quantitative Relationships in Extant Alligators”. From Tyrannosaurid Paleobiology, edited by J. Michael Parrish, Ralph E. Molnar, Philip J. Currie, and Eva B. Koppelhus. Bloomington: Indiana University Press, 2013. Pages 134-154. https://people.ohio.edu/witmerl/Downloads/2013_Hurlburt_Ridgely_&_Witmer_tyrannosaurid_relative_brainsize.pdf.
- Li, Rihui; Lockley, Martin G.; Makovicky, Peter J.; Matsukawa, Masaki; Norell, Mark A.; Harris, Jerald D.; Liu, Mingwe (2008). “Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China”. Naturwissenschaften, volume 95, issue 3 (April 2008). Pages 185-191. http://cactus.dixie.edu/jharris/Dromaeopodus.pdf.
- Manning, Philip L.; Payne, David; Pennicott, John; Barrett, Paul M.; Ennos Roland A. (2006). “Dinosaur killer claws or climbing crampons?”. Biology Letters, volume 2, issue 1 (March 22, 2006). Pages 110-112.
- Orellania, Sergio Alvarado; Figueroa Rojas, Ricardo A. (2005). “Possible social foraging behavior in the Red-Backed Hawk (Buteo polyosoma)”. Ornitologia Neotropical, volume 16 (2005). Pages 271-275. https://www.academia.edu/3513923/Possible_social_foraging_behavior_in_the_Red-backed_Hawk_Buteo_polyosoma_.
- Roach, Brian T.; Brinkman, Daniel L. (2007). “A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs”. Bulletin of the Peabody Museum of Natural History, volume 48, issue 1 (April 2007). Pages 103-138. http://pds17.egloos.com/pds/201004/29/62/Theropod_Dinosaur.pdf.
Websites:
- Beyond Bones – Blog of the Houston Museum of Natural Science. “Raptors – Group Hunters or Cannibals?”, by Robert Bakker (March 11, 2010). https://blog.hmns.org/2010/03/raptors-group-hunters-or-cannibals/.
- Fossil Crates. “Pack Hunting Raptors – What is the evidence?”, by Dr. Brian Curtice (May 21, 2020). https://www.fossilcrates.com/blogs/news/pack-hunting-raptors-what-is-the-evidence.
- Mark Witton Blogspot. “Realistic raptors: pop-culture dromaeosaurs vs current science, part 1”, by Mark Witton (March 27, 2020). http://markwitton-com.blogspot.com/2020/03/realistic-raptors-pop-culture.html.
- National Geographic. “Deinonychus and Velociraptor used their killing claws to pin prey, like eagles and hawks”, by Ed Yong (December 14, 2011). https://www.nationalgeographic.com/science/article/deinonychus-and-velociraptor-used-their-killing-claws-to-pin-prey-like-eagles-and-hawks.
- National Geographic. “Troodon sapiens?: Thoughts on the ‘Dinosauroid’”, by Riley Black (October 23, 2007). https://www.nationalgeographic.com/science/article/troodon-sapiens-thoughts-on-the-dinosauroid.
- Natural History Magazine. “Day of the Deinos: Did predatory dinosaurs leave clues to their pack-hunting habits at kill sites?” by Desmond Maxwell (December 1999-January 2000). https://www.naturalhistorymag.com/htmlsite/master.html?https://www.naturalhistorymag.com/htmlsite/1299/1299_feature.html.
Videos (listed in order of air date):
- Dinosaurs, Dinosaurs, Dinosaurs. Hosted by Gary Owens and Eric Boardman. Twin Tower Enterprises, 1985.
- Dinosaur! Episode 1 – “The Tale of a Tooth”. Hosted by Walter Cronkite. A&E, 1991.
- Dinosaur! Episode 3 – “The Tale of an Egg”. Hosted by Walter Cronkite. A&E, 1991.
- Paleoworld. Season 2, episode 11 – “Troodon: Dinosaur Genius”. The Learning Channel, 1995.
- Paleoworld. Season 4, episode 7 – “Killer Raptors”. The Learning Channel, 1997.
- Walking with Dinosaurs: The Ballad of Big Al. BBC, 2001.
- Dino Lab. The Discovery Channel, 2006.
Categories: Paleontology, Uncategorized

Sorry to be the “actually” guy, but the “Pack Hunters” & “Intelligence” sections are a bit problematic, the former due to an over-reliance on very flawed sources: Roach/Brinkman 2007, which is very flawed for reasons discussed elsewhere (See #2: https://www.deviantart.com/jd-man/journal/SD-Top-4-most-annoyingly-popular-dino-hypotheses-395469447 ); Frederickson et al. 2020, which is very flawed for reasons discussed elsewhere (See “New #1 Contender”: https://www.deviantart.com/jd-man/journal/SD-Most-annoyingly-popular-dino-hypotheses-addend-842963661 ); Curtice’s blog post, which is over-reliant on Roach/Brinkman 2007 & Frederickson et al. 2020. The following problems are especially notable:
-1) You gloss over 1 of the most important pieces of evidence for pack-hunting Deinonychus (“Typically, paleontologists find three or four Deinonychus teeth associated with Tenontosaurus bones”). To quote Maxwell/Ostrom, “Deinonychus, with approximately 70 teeth, would have had to replace its teeth at an unrealistically rapid rate if it were to lose two or three teeth per meal, as such drastic tooth loss would leave the animal toothless after only about 20 meals” ( https://sci-hub.st/https://www.jstor.org/stable/4523664 ). In other words, Deinonychus/Tenontosaurus associations typically represent at least 2 (probably 3 or 4) Deinonychus.
-2) In reference to MOR 682, you mistakenly claim that the 11 Deinonychus teeth were “scattered around” the Tenontosaurus skeleton. In actuality, they “were found in the region of the abdomen and pelvis, suggesting that the predators lost their teeth while feeding on the viscera. Most modern carnivores begin with the areas around the anus and abdomen when they feast on freshly killed prey, and it’s likely that carnivorous dinosaurs did the same[…]This may indicate that the pack fed on the abdominal contents while they were still warm and moist. If[…]the carcass was scavenged over time by many individuals[…E.g. A non-cooperative mob…]we would expect a much more disturbed carcass and a wider scattering of teeth. Similarly, if the Tenontosaurus had been killed by a larger predator[…]the carcass would have been strewn around the area” ( https://www.naturalhistorymag.com/htmlsite/master.html?https://www.naturalhistorymag.com/htmlsite/1299/1299_feature.html ). As pointed out by Maxwell/Ostrom above, the same goes for Deinonychus/Tenontosaurus associations in general (See page 710 under “Discussion”).
-3) In reference to Li et al. 2007, you forget to mention that it shows how misleading/wrong Roach/Brinkman’s trackway argument is (“Roach and Brinkman[…]proposed that trackway data previously presented in support of gregarious nonavian theropods are perhaps better interpreted as coincidental instances of normally solitary individuals converging on a common point”: http://cactus.dixie.edu/jharris/Dromaeopodus.pdf ).
-4) Also in reference to Li et al. 2007, “it is clear that the animals were not hunting at the time” ( http://scienceblogs.com/laelaps/2007/10/30/at-long-last-dromeosaur-tracks/ ). To quote Bakker, “predators don’t usually hang out in groups if they don’t hunt together. Tigers are like this — they mostly hunt alone, and you don’t see bunches of tigers lying around together. But lions are social predators. They hunt and raise their young and sleep and snore together” (See “Raptor Pack”). The “raise their young[…]together” part is especially important b/c “cooperative hunting[…]is generally related to cooperative breeding” ( https://www.academia.edu/3513923/Possible_social_foraging_behavior_in_the_Red-backed_Hawk_Buteo_polyosoma_ ). Li et al. 2007 seems to agree (“The discovery of six parallel, closely spaced D. shandongensis trackways provides compelling, independent evidence for at least occasional[…]“pack” or family group[…]behavior in the track-making animals, comparable to what has been demonstrated in other dinosaurs”).
-5) In reference to the “Intelligence” section: 1st, you mistakenly claim “that the raptors possessed brains which were only slightly more advanced than” other theropods; As indicated by Buchholtz 2012 & the sources cited therein (See the Buchholtz quotes below, which are from the 2nd edition of “The Complete Dinosaur”), non-bird maniraptoran brains were “strikingly different” from tyrannosaurid & allosauroid brains; 2nd, you mistakenly claim that opossums “are certainly not possessed with remarkable intelligence”; In actuality, “results from some learning and discrimination tests rank opossums above dogs and more or less on a par with pigs in intelligence (Hunsaker and Shupe 1977)” ( https://books.google.com/books?id=-xQalfqP7BcC&pg=PA25&dq=%22tests+rank+opossums%22 ). 3rd, you mistakenly refer to chickens as “dumb”; In actuality, chickens 1) “probably fall about mid-range on the intelligence scale of birds” ( https://books.google.com/books?id=Ct4-qGkuC-kC&pg=PA34&dq=%22fall+about+mid-+range%22&hl=en&sa=X&ved=0ahUKEwjw-cjpgOvOAhXKJx4KHS2lCx4Q6AEIKjAC#v=onepage&q=%22fall%20about%20mid-%20range%22&f=false ), & 2) “are intelligent animals, outperforming dogs and cats on many tests of advanced cognition. As just one example, in a study by the Silsoe Research Institute in England, researchers showed that chickens have the ability to make a conscious choice to delay gratification” ( https://issuu.com/grantjustinflowers/docs/annual_report_book ); Said study is Abeyesinghe et al. 2004 ( https://www.silsoeresearch.org.uk/animal-welfare/siobhan/scarticle.pdf ); To be fair, it’s Larsson’s fault for not getting his facts straight before making such claims; However, in the future, I recommend verifying such claims (including the opossum one) independently.
Quoting Buchholtz: “Brain anatomy in theropods is relatively well known and falls into two distinct anatomical groups, suggesting different dominant sensory modalities and lifestyles. Allosaurids and large coelurosaurs retain primitive endocast shape and distinct cerebral and pontine flexures (Fig. 10.2). The optic tectum is obscured and must have been small. The cerebral hemispheres are not separable and show only modest convexity. The prediction of extremely large olfactory bulbs in Tyrannosaurus rex (Brochu 2000) has been scaled downward, but it still seems likely that smell was a dominant sense (Rogers 1998, 1999; Stokstad 2005; Saveliev and Alifanov 2007). Rogers (1998, 1999) described the vestibular apparatus of Allosaurus fragilis, which more closely resembles that of crocodylians than that of lizards, turtles, or birds. He used semicircular canal orientation to suggest that Allosaurus held its head at or very slightly (≤10 degrees) inclined to the horizontal. Burish et al. (2004) included endocranial casts of Tyrannosaurus, Allosaurus, and Carcharodontosaurus in an analysis of relative telencephalic (~cerebral) size and behavior in birds. Their data indicate values for theropods at or below the range of living reptiles, and predict low social complexity.”
Quoting Buchholtz: “Endocasts of small coelurosaurs (Russell 1969, 1972; Colbert and Russell 1969; Kundrát 2007) display a strikingly different anatomical pattern (Fig. 10.6). They retain details of brain anatomy and roofing bone sutures on their surfaces, suggesting that the brain filled the braincase nearly completely. Brain flexures are minimal and olfactory bulbs are small, indicating that smell was not a dominant sense. Cerebral hemispheres are separable, convex, and expanded laterally and/or posteriorly (Kundrát 2007), suggesting an active intelligence. The large optic lobes are visible either dorsally or displaced laterally by the large cerebrum, as in living birds. Russell (1969) associated the large optic lobes with large eyes and binocular vision, and it is likely that sight was the dominant sense. Kundrát (2007) described an expanded cerebellum with presumptive cerebellar folia among the avianlike characters of the oviraptorid theropod Conchoraptor, inferring excellent balance and coordination.
Encephalization quotients of small coelurosaurs vary with predictions of body mass and percentage of braincase fill, but even conservatively, they are far higher than those of any other dinosaur group, overlapping those of living birds (Hopson 1977; Kundrát 2007). Larsson et al. (2000) estimated cerebral volumes by superimposing ellipsoids on endocasts with surficial indications of cerebral extent. Their data suggest at least three stages of increase of relative cerebral size to total brain size over a period of only 40 million years: of coelurosaurs over allosaurs, of Archaeopteryx over coelurosaurs, and of ornithurine birds over Archaeopteryx. The high encephalization values of small coelurosaurs indicate an active, complex, and social lifestyle that agrees well with their frequent interpretation as pack hunters.”
There’s no need to apologize for being the “actually” guy. Just know that I based this article upon the information which was available to me at the time that I was researching and writing it, and there were scientific publications concerning this topic which I simply did not have access to.
I noticed in your first criticism that you only half-quoted me. The full quote reads “Typically, paleontologists find three or four Deinonychus teeth associated with Tenontosaurus bones. However in 1992, a sub-adult Tenontosaurus skeleton measuring 14 feet long was found, and scattered around it were eleven Deinonychus teeth of varying sizes. It would be very debilitating if any single Deinonychus lost eleven of its teeth in one instance, so the only conclusion is that these teeth came from the mouths of multiple Deinonychus which were feeding upon the corpse at the same time”. So you see, I never stated or implied that Deinonychus didn’t congregate at kills. However just because multiple individuals were feeding at a kill site, that doesn’t necessarily mean that they all belonged to a single social group. Bakker’s argument concerning predators and social behavior, which you gave in your comment, is very limiting. Predators often do congregate, despite not being part of an actual social group. Komodo dragons, crocodilians, grizzly bears, some birds-of-prey, and some species of sharks do this regularly. Regarding MOR 682, the location of the shed Deinonychus teeth around the belly of the sub-adult Tenontosaurus skeleton demonstrates that these individuals were feeding on the same area of the carcass, but it does not demonstrate that they were all part of a single group. Carnivores will naturally attack the soft parts of a dead animal first since that’s where the most meat is and they don’t want to expend energy unnecessarily by crunching through bones. Fights could have broken out between the various carnivores over the choicest portions, which would explain the bite marks and claw marks which have been on Deinonychus bones which were likely made by other members of its kind as they scrapped with each other over getting their portion of meat.
I did not have access to Li et al (2007) at the time that I was researching and writing this article, and I based my own article upon the information which was available to me during that time; thank you for providing a PDF link to it (http://cactus.dixie.edu/jharris/Dromaeopodus.pdf). Li et al essentially argued that since multiple trackways which were apparently made by the same species of animal were all moving in the same direction, that was strong evidence of gregariousness. The best assertions for group behavior are found on pages 189-190, in which the authors state “The tracks do not overlap, footprint size is almost uniform between trackways, and ‘intertrackway’ spacing (Lockley 1989) is regular and less than a single stride length, all of which strongly suggest that they were registered by a group of six individuals moving together as a coherent group, regardless of possible physiographic ‘control’. Because the sedimentology indicates a relatively rapid emplacement and subsequent burial of the trackways, a scenario in which six similarly sized individuals independently generated parallel, regularly spaced, nonoverlapping trackways is highly unlikely. Whether such an aggregation of similarly sized individuals represents a common or rare (or perhaps seasonal) event cannot be determined from a single find, but it indicates that dromaeosaurids in fact engaged in some type of group behavior”.
However, while the evidence as they present it is compelling, I think that the researchers may have been jumping to conclusions. I must point out that many of these “trackways” (which are visible on the bottom of page 188) are incomplete, and four of them consist of just a single footprint. A single footprint is not a trackway, it’s just a footprint. Saying that none of the trackways overlap, while true in the strictest sense, is quite presumptuous, as the researchers would have no idea if these trackways overlapped based upon just a single footprint from each trackway.
As I wrote, “Just because a footprint trackway was discovered showing several individuals moving in the same direction, that does not mean that they were all moving together in a single group, and it does not mean that each of these individual trackways were all made at exactly the same time. Minutes or hours may have passed between the times that each individual animal came that way”.
The fact that there are trackways of at least three different species of animals, some moving in one direction and others moving in the opposite direction (as shown by the image on page 188 of the Li et al article), and all of these trackways seem to have been laid down within a short time (which, in this case, might be throughout a 24 hour period or perhaps even longer) lead me to suspect that these sets of footprints might actually be evidence of a prehistoric “game trail”, in which wild animals will travel along a certain pathway either due to obstructions in the landscape which prevents them from moving elsewhere or even out of prolonged habit. It has been known for some time that herding animals such as deer, bison, hippos, and elephants will frequently use the exact same routes so often that they eventually become well-worn pathways. Carnivores such as wolves and mountain lions eventually figure this out, and will either lurk in ambush along these prey highways, waiting for their prey to come down the path, or travel up and down these routes themselves searching for prey. On page 189, the authors allude to this possibility when they refer to the terrain that these footprint trackways were made as a “physically controlled” landscape, which constrains animals to move only within a certain confined area. In this case, the authors also state that the terrain in which these footprints were made was composed of sandstone and featuring mud drape and ripple marks, and that the sedimentology of the site indicates it was the shoreline of a river. The authors also stated “The absence of desiccation cracks indicates the footprints were emplaced within a very short time span”, but if you’re walking along the muddy sandy shore of a river, it’s very unlikely that the soil is going to dry out unless with a massive reduction of the water level, which evidently did not happen here.
It is true that maniraptoran theropods did possess more advanced brains than non-maniraptorans – I am not denying that. Understandably, earlier theropods like megalosaurs and carnosaurs possessed primitive crocodilian-like brains, whereas advanced theropods like ornithomimids, oviraptorosaurs, and especially raptors, which were more closely related to birds than to crocodiles, possessed more bird-like brains. Dale Russell also plotted the encephalization quotient among raptor dinosaurs and concluded that raptors like Troodon were placed higher in intelligence than other dinosaurs. However, considering how “smart” other dinosaurs were, being only slightly smarter than them was not much of an improvement. Raptors were certainly not as smart as many modern mammals, and they certainly would not have had advanced problem-solving abilities like the media commonly states. As I said earlier, the intelligence of raptors, and Troodon especially, has been very over-exaggerated. His assessment was that Troodon was about as smart as the primitive mammals which were around then, like the opossum-like marsupials which dwelt in the Mesozoic forests.
Speaking of opossums, concerning your comment about opossum intelligence, experiments conducted throughout the late 1950s by Dr. William T. James (1903-1988) showed that wild-caught opossums were unable to develop conditioned responses to stimuli, even after running the trials 300 times. However, captive-raised opossums were much more responsive and were able to complete tasks far quicker, possibly due to their prolonged association with humans, and exhibited a marked improvement in intelligence and complex behavior in comparison to their wild relatives. Other methods which employed electro-shock (which I definitely do not approve of) showed more promising results. Experiments involving mazes and operating simple mechanical devices showed that opossums were certainly capable of performing these tasks, but it must also be noted that James had transitioned from wild opossums to lab-raised opossums because the wild opossums had proven too unresponsive. Lab-raised opossums, who had spent their entire lives around humans and were presumably constantly exposed to new and varied stimuli were given more latitude for their brains to develop. In essence, James skewed his results by using animals which were uncharacteristically smarter than their wild-dwelling relatives, and that was how he as able to gain the results that he got. That would be like if aliens were determining how smart your average human was by having Albert Einstein or Stephen Hawking complete a math test, and then claiming that all humans must be as smart as that (Don Hunsaker II, ed., “The Biology of Marsupials”. New York: Academic Press, 1977. Pages 333-335).
Finally, I never said that chickens were dumb. Hans Larrson said that Troodon was about as smart as a modern-day chicken, and not just any old chicken, but one which was especially not too bright.
“Just know that I based this article upon the information which was available to me at the time that I was researching and writing it, and there were scientific publications concerning this topic which I simply did not have access to.”
No worries, I figured that might’ve been the case, but didn’t wanna assume. Hopefully, this reply clears things up.
“I noticed in your first criticism that you only half-quoted me.”
I only quoted the parts that seemed especially problematic. Sorry if that wasn’t clear. In this case, my point was that, while Deinonychus/Tenontosaurus associations typically represent multiple Deinonychus, you only discussed 1 example as possible evidence for pack-hunting Deinonychus (MOR 682).
“Bakker’s argument concerning predators and social behavior[…]is very limiting. Predators often do congregate, despite not being part of an actual social group. Komodo dragons, crocodilians, grizzly bears, some birds-of-prey, and some species of sharks do this regularly.”
I think you’re missing the point: If the Li et al. 2007 trackmakers were hunting, you could argue that they’re multiple lone hunters independently converging on the prey; However, as I pointed out in my 1st comment, said trackmakers clearly weren’t hunting; As Bakker pointed out, you don’t usually see bunches of lone hunters like tigers hanging out/lying around/raising their young/sleeping & snoring together the way you do pack hunters like lions; I’m not saying it’s impossible that the Li et al. trackway represents something unusual, just that regardless of your nit-picks, the circumstances of said trackway are more suggestive of pack hunters than multiple lone hunters.
“Regarding MOR 682, the location of the shed Deinonychus teeth around the belly of the sub-adult Tenontosaurus skeleton demonstrates that these individuals were feeding on the same area of the carcass, but it does not demonstrate that they were all part of a single group.”
I think you’re missing the point: See pages 271-2 of Orellana/Rojas 2005 (link in my 1st comment, #4); Among pack hunters, “prey are shared according to some social order”, hence why (to quote Maxwell) “we have a concentration of teeth around the abdomen and pelvis”; Among other social foragers, “prey are chaotically shared or not shared at all”, hence why (to quote Maxwell) “we would expect a much more disturbed carcass and a wider scattering of teeth.” Basically, the circumstances of the Deinonychus/Tenontosaurus associations are more suggestive of fresh kills by cooperative packs than scavenging events by non-cooperative mobs.
“It is true that maniraptoran theropods did possess more advanced brains than non-maniraptorans – I am not denying that[…]However, considering how “smart” other dinosaurs were, being only slightly smarter than them was not much of an improvement. Raptors were certainly not as smart as many modern mammals,”
1stly, no offense, but you seem to be contradicting yourself: As I pointed out in my 1st comment, non-bird maniraptoran brains were “strikingly different” from tyrannosaurid & allosauroid brains; You say you understand, but then repeat the misleading/wrong claim that the former were “only slightly smarter” than the latter; No offense to crocs, but last I checked, there’s a big difference btwn average croc intelligence & average bird intelligence ( https://www.youtube.com/watch?v=iZHBhDT2MZ8 ).
2ndly, it depends on what you mean by “many modern mammals”: As I pointed out in my 1st comment, chickens “are intelligent animals, outperforming dogs and cats on many tests of advanced cognition”. Based on what I’ve read, Archaeopteryx were similarly intelligent to chickens (As you may remember, this paper is cited in Buchholtz 2012: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.583.8968&rep=rep1&type=pdf ), & non-bird maniraptorans were similarly intelligent to or more intelligent than both (See the 1 short Walsh/Knoll quote below. Hopefully, it makes my original point clearer than the 2 longer Buchholtz quotes in my 1st comment).
“Speaking of opossums,[…]That would be like if aliens were determining how smart your average human was by having Albert Einstein or Stephen Hawking complete a math test, and then claiming that all humans must be as smart as that”
Based on what I’ve read, a better analogy would be to compare Dr. William T. James’s experiments to “determining how smart your average human was by[…waking random ppl up in the middle of the night to…]complete a math test”. Put another way, “it must be remembered that the opossum is a nocturnal species. Many of the published observations and stories concerned with the behavior of the opossum were made during the daylight hours at a time when the opossum is normally asleep. If one considers the time of observation it is quite understandable that the opossum gained the reputation of being a bit sluggish in some of the early accounts” (See pages 31-2 AWA the sources listed at the end of this book for more info: https://www.researchgate.net/publication/265347494_The_Opossum_Its_Amazing_Story ).
“I never said that chickens were dumb. Hans Larrson said that Troodon was about as smart as a modern-day chicken, and not just any old chicken, but one which was especially not too bright.”
Fair enough. However, when quoting/paraphrasing ppl in the future, you should use quotations/brackets. Not doing so implies that the statement is in your own words. In this case, Larrson wrongly claimed that “the Troodon[…]is not so different from an unintelligent bird[…]something like a chicken”, & the narrator followed up w/the misleading claim that “even the intelligence of a dumb chicken was an advantage that gave Troodon an important edge in prehistoric times” ( https://www.youtube.com/watch?v=Uu4orxfYW_s ). In the future, you should also avoid using very flawed dino docs like “Dino Lab” as sources (E.g. Despite having access to actual Troodon animation, DL uses Coelophysis animation & calls it Troodon).
Quoting Walsh/Knoll ( https://www.academia.edu/1061233/Directions_in_Palaeoneurology ): “It is now clear that while some theropod clades pos-sessed typically ‘reptile’-like brains (Giffin et al. 1988; Gif-fin 1989; Rogers 1999; Brochu 2000; Franzosa and Rowe 2005; Sanders and Smith 2005; Witmer and Ridgely 2009), at least some maniraptoran theropods had surpris-ingly bird-like brains (Kundra´t 2007; Balanoff et al. 2009; Norell et al. 2009). In these taxa, not only is the telen-cephalon significantly enlarged (sometimes to an extent greater than in Archaeopteryx), but flight-related regions such as the cerebellar flocculus, which are especially enlarged in birds, are also very well developed (Kundra´t 2007). This has fuelled the debate about whether these taxa, which often bear feathers, are in fact secondarily flightless birds rather than bird-like theropods (Witmer 2009; Kavanau 2010).”
Thank you for clarifying certain things which were a bit confusing to me. Consequently, I have updated the article based upon this new information and the interpretations of that information.